 Reproduced, with permission, from:
Häder, D. P., R. C. Worrest, and H. D. Kumar. 1991. Aquatic ecosystems. Chapter 4 in Environmental effects of ozone depletion: 1991 update. Nairobi: United Nations Environment Programme.
Reproduced, with permission, from:
Häder, D. P., R. C. Worrest, and H. D. Kumar. 1991. Aquatic ecosystems. Chapter 4 in Environmental effects of ozone depletion: 1991 update. Nairobi: United Nations Environment Programme.
Panel Report Pursuant to Article 6 of the Montreal Protocol on Substances that Deplete the Ozone Layer Under the Auspices of the United Nations Environment Programme (UNEP)
November 1991
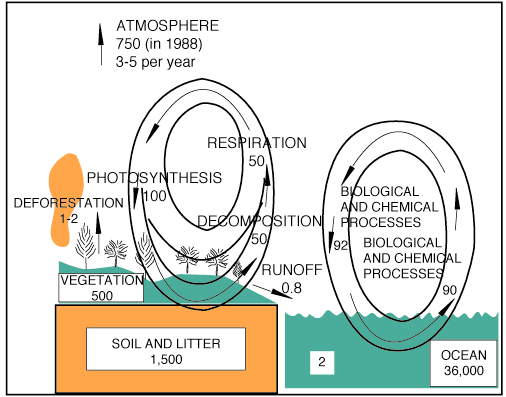
Aquatic ecosystems contribute more biomass (104 Gt/a) than all terrestrial ecosystems (100 Gt/a) combined. Recent work on UV-B effects has concentrated on inhibition mechanisms and field studies in the subpolar waters of Antarctica, because of its high productivity and the occurrence of the ozone hole over this region.
Phytoplankton organisms orient within the water column using external factors. However, mobility and orientation mechanisms are impaired by UV-B radiation. Because most organisms do not possess UV-B receptors, they cannot avoid deleterious wavelength radiation that (according to new measurements) penetrates deeper into the water column than what has been previously measured. New action spectra indicate that, in addition to DNA, other targets absorb UV-B radiation including intrinsic proteins of the photoreceptor and photosynthetic apparatus.
The inability to adjust their position within the water column causes massive inhibition of photosynthesis, measured both in field and laboratory studies. Only in a few cases have potential UV-B-inducible screening pigments been identified.
A large share of the nitrogen consumed by higher plants is made available by bacterial microorganisms, which have been found to be very sensitive to UV-B radiation. Losses in nitrogen fixation could be compensated by additional nitrogen fertilization. However, such actions could stress the capabilities of developing nations.
The role of DMS, released from plankton and macroalgae as aerosol and cloud nuclei, is of major concern. Most importantly, a UV-B-induced decrease in phytoplankton populations may have an impact on cloud patterns and concomitant global climate changes.
An increased understanding of Antarctic trophic dynamics suggests that the likelihood of direct UV-B radiation effects on consumers is small. Rather, it is the possibility of indirect effects that may significantly affect the Antarctic trophic structure, such as different species sensitivities to UV-B radiation, or decreases in total available primary production. Because more than 30% of the world's animal protein for human consumption comes from the sea, the human populations may also be affected by the direct and indirect consequences of increased solar UV-B radiation on aquatic food webs.
Another potential consequence of a decrease in marine primary productivity would be a reduction in the capacity of the ocean to absorb carbon dioxide. A hypothetical loss of 10% of the marine phytoplankton would reduce the oceanic annual uptake of carbon dioxide by about 5 Gt (an amount equal to the annual emissions of carbon dioxide from fossil fuel consumption). Uncertainties regarding the magnitude of increased levels of UV-B radiation on aquatic systems still remain, including problems of extrapolating laboratory findings to the open sea, and the nearly complete absence of data on long-term effects and ecosystem responses. Uncertainties and future research needs include adaptive strategies and the effects of cumulative UV-B radiative doses. Additional information is needed in several areas before a more reliable assessment of risks is possible.
Current data suggest that predicted increases in UV-B radiation could have important negative effects on the marine environment. However, uncertainties regarding the magnitude of these effects still remain, including problems of extrapolating laboratory findings to the open ocean, and the nearly complete absence of data on long-term effects and ecosystem responses.
Planktonic marine organisms account for over half of the total global amount of carbon fixed annually (10[11] tons). Any reduction in this productivity will undoubtedly affect global food supply and global climate. Both primary production and subsequent steps in biological food webs are sensitive to current UV-B levels, and are potentially endangered by expected increases in UV-B radiation.
UV-B radiation affects 1) adaptive strategies (e.g., motility, orientation), 2) impairs important physiological functions, (i.e., photosynthesis and enzymatic reactions), and 3) threatens marine organisms during their developmental stages (e.g., the young of finfish, shrimp larvae, crab larvae). In addition to DNA damage, UV-B radiation affects enzymes and other proteins, eliciting photodynamic responses. These effects can have a number of possible consequences for aquatic ecosystems:
However, additional information is needed before more reliable assessments of the UV-B radiative risks posed to the marine environment will be possible.
Since the release of the UNEP Environmental Effects Panel Report of 1989 [UNEP, 1989], UV-B aquatic research has concentrated on phytoplankton and the Antarctic ecosystem. It is estimated that phytoplankton convert 104 billion tons of carbon into organic material annually, which is slightly more than the 100 billion tons from all terrestrial ecosystems combined [Houghton and Woodwell, 1989] (Figure 4.1). Phytoplankton, at the base of the aquatic food chain/trophic structure, serve as food for primary consumers (e.g., larvae of fish and shrimp), which in turn are consumed by secondary and tertiary consumers (e.g., fish). The final consumers of this trophic web are large fish, birds, and mammals, including man. Thus, any changes in the size and composition of phytoplankton communities will directly affect man's marine food sources.
A decrease in springtime concentrations of stratospheric ozone of over 50% have been clearly documented in the Antarctic. Within the Antarctic ecosystem, biomass is concentrated as an aquatic component, as opposed to the terrestrial environment. Concentrations of phytoplankton in subpolar waters may be 10[3] to 10[4] times greater than concentrations of phytoplankton found in tropical and subtropical seas [Jeffery and Humphrey, 1975]. Any significant increase in UV-B radiation, due to thinning of the ozone layer, could well diminish growth and productivity of phytoplankton, subsequently affecting all higher trophic levels in the aquatic food web. Therefore, it is not surprising that a majority of recent research has looked at the effects of increased UV-B radiation in Antarctic waters. Ongoing research activities include investigations of both direct (physiological and behavioral) and indirect effects (trophic implications).
Because of their requirement for solar energy, phytoplankton dwell in the top layers of the water column, the photic zone [Ignatiades, 1990]. Their position within the column is maintained by precise orientation strategies using light, gravity, and other external factors as guides. Phytoplankton, in the photic zone, would be exposed to any increase in solar ultraviolet radiation. Most phytoplankton organisms (studied up to now) do not have UV-B photoreceptors to guide them away from harmful radiation, a situation similar to humans. Previous work demonstrated that mobility/orientation mechanisms in response to light are impaired by solar ultraviolet radiation [Häder and Worrest, 1991] penetrating deep into the photic zone [Baker and Smith, 1982]. The ability of phytoplankton to adjust their position within the water column, in response to constantly changing conditions, may be affected at even ambient UV-B levels. In fact, ambient UV-B fluxes may cause damage to some species of phytoplankton. However, it should be emphasized that there are uncertainties regarding the magnitude of these effects, including problems of extrapolating laboratory findings to the open sea, and the nearly complete absence of data on long-term effects and ecosystem responses. Likewise, there is a need to investigate adaptation mechanisms. Before effects of exposure to solar UV-B radiation can be predicted, information is required on seasonal abundances and vertical distributions of marine organisms, vertical mixing, and the penetration of UV-B radiation into appropriate water columns.
In their natural habitats, organisms are exposed to a wide range of UV-B radiative doses. This radiation has been shown to affect growth, photosynthesis, nitrogen incorporation, and enzyme activity [Döhler and Alt, 1989; Döhler, 1990]. Even a small increase in global UV-B radiation would intensify maximal irradiation levels, as well as increase long-term exposure doses.
Primary Producers
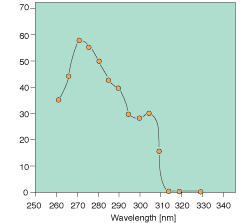
Recent results indicate that orientation mechanisms responsive to both light and gravity are affected by solar ultraviolet radiation in a number of ecologically significant phytoplankton groups [Häder and Liu, 1991]. Action spectra inhibitory effects are different from the DNA absorption spectrum and the action spectra calculated for higher plants, indicating that UV radiation affects these organisms by a totally different mechanism (Figure 4.2). Recent data indicate that proteins essential for specific functions in orientation and photosynthesis are the primary targets of UV-B radiation.
Biochemical analyses, conducted to reveal the molecular targets of UV-B inhibition, show that specific photoreceptor proteins are degraded by ultraviolet radiation. Simultaneously, photosynthetic pigments (responsible for converting solar energy) are bleached and destroyed by radiation. The results of these biochemical studies are further supported by spectroscopic investigations showing losses in pigmentation.
In order to evaluate the effects of enhanced ultraviolet radiation, the vertical movement of natural phytoplankton was analyzed in 3-m long Plexiglas columns [Eggersdorfer and Häder, 1991]. Most organisms moved to the surface during daytime hours, although some species avoided periods of intense ultraviolet radiation during the midday hours by moving slightly down in the water column. However, this avoidance response is not sufficient to protect organisms under conditions of increased UV-B irradiation [UNEP, 1989; Yoder and Bishop, 1990].
The UV-B irradiance in Antarctic waters drastically increased during the occurrence of the ozone hole [Bidigare, 1989; Lubin et al., 1989; Karentz and Lutze, 1990]. However, changes in marine productivity accompanying UV flux changes have not been determined. Recent measurements show that UV-B penetrates 65 meters deep in clear Antarctic waters [Gieskes and Kraay, 1990]. Consequently, measurements of photosynthetic biomass production in Antarctic waters under the ozone hole show a pronounced decrease of productivity by up to 25% [Holm-Hansen, 1990]. ATP content of Antarctic phytoplankton cells (a reflection of energetic state), significantly decreased in the presence of the ozone hole [Vosjan et al., 1990; Karentz et al., 1991a].
Field studies indicate that photosynthesis is impaired first, followed by decreases in protein concentration and changes in pigment composition. As a result, a dramatic decrease in photosynthetic oxygen production can be measured after exposure to solar radiation [Smith et al., 1980; Zündorf and Häder, 1991]. It should be mentioned that other spectral bands, such as UV-A and visible radiation, may contribute to photosynthetic inhibition [Smith et al., 1980; Mitchell, 1990]. Likewise, photosynthetic inhibition has been detected in macroalgae at their natural depth [Bittersmann et al., 1988; Nultsch et al., 1990].
Higher plants are known to produce UV-B absorbing substances when exposed to ultraviolet radiation, thus protecting them from solar short-wavelength radiation [UNEP, 1989]. Recently, UV-absorbing substances have been isolated from phytoplankton, as well as from a number of primary and secondary feeders [Carretto et al., 1990; Karentz et al., 1991b]. However, all of these mycosporine-like amino acids have maxima in the UV-A range and only secondary peaks in the UV-B. In addition, it is not clear whether the production of these potentially screening substances can be induced by exposing organisms to ultraviolet radiation [Raven, 1991]. One exception is cyanobacteria where a UV-B inducible pigment has been found within the slime sheath surrounding the organisms, which absorbs up to 88% of the UV-B radiation [Garcia-Pichel and Castenholz, 1991].
Some planktonic bacteria have the capability to incorporate atmospheric nitrogen into a form accessible to higher plants. Higher plants cannot assimilate atmospheric nitrogen themselves and are therefore restricted to using nitrogen in the form of nitrate, nitrite, or ammonia. A large share of the nitrogen consumed by higher plants is made available by bacterial microorganisms, (e.g., in tropical rice paddies) [Kumar and Kumar, 1988]. However, these microorganisms are highly sensitive to solar UV-B radiation. Consequently, losses in nitrogen fixation due to increases in UV-B radiation may need to be compensated for by artificial nitrogen fertilization. Cyanobacteria alone are estimated to produce 35 million tons of nitrogen annually.
Furthermore, many algal groups are known to produce dimethylsulphonio-propionate as an antifreeze and osmotic regulator. From this substance, dimethylsulphide (DMS) is cleaved enzymatically and emitted into the surrounding water [Karsten et al., 1990]. From there it is released into the atmosphere where it undergoes photochemical oxidation to sulphate and methane sulphonate. These substances are thought to act as nuclei in aerosol formation and cloud condensation. Forty million tons of sulphur are released by this mechanism annually, equaling about 50% of the sulphur emissions from anthropogenic sources. Therefore, there are concerns that a reduction in the phytoplankton could have an effect on global climate changes [Andreae, 1986].
Recently, little work has been conducted on the direct effects of UV-B radiation on zooplankton. Over the last few years, however, a greater understanding of Antarctic trophic dynamics has developed. Furthermore, there is a growing appreciation for microorganisms (especially diatoms) found in the sea ice [Garrison et al., 1986]. This community may be a significant factor for productivity in Antarctic waters, contributing to biomass and serving as an inoculum for spring phytoplankton blooms [Voytek, 1990].
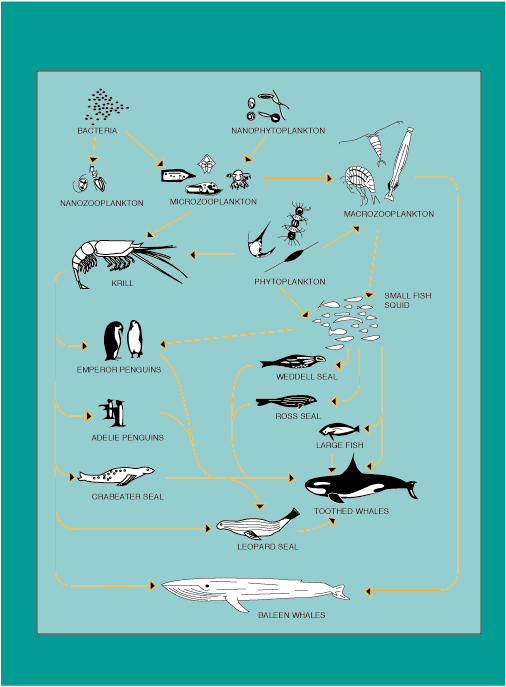
Figure 4.3 illustrates the nature of the Antarctic aquatic food web. Voytek [1990] suggests that it is inadequate to look only at primary production, but rather, the effects of increased UV-B radiation on each trophic component must be investigated. In general, direct effects from radiation on consumers is small, however, indirect effects (such as changes in the composition of phytoplankton assemblages) may significantly affect the Antarctic trophic structure. This effect is the result of species-specific sensitivity to UV-B radiation or decreases in total available primary production.
Human populations may also be affected by direct and indirect consequences of increased solar UV-B radiation on aquatic food webs. Because more than 30% of the world's animal protein for human consumption comes from the sea (in many developing countries this percentage is even larger), a substantial decrease in biomass production would diminish fishery resources in the face of growing world populations.
Another important consequence of a potential decrease in marine phytoplankton productivity would be the reduced ability of the ocean to serve as a sink for atmospheric carbon dioxide. Phytoplankton not only produce more than half of the biomass on our planet, but also absorb and fix more than half of the carbon dioxide from the atmosphere. Hypothetically, a loss of only 10% of the phytoplankton would prevent about 5 gigatons of carbon (in the form of carbon dioxide) from being removed from the atmosphere annually (which is equal to the amount of carbon dioxide emitted currently by fossil fuel utilization). Ten to twenty percent of the gross primary production from the oceans is estimated to occur in southern regions [Voytek, 1990]. Because carbon dioxide is a greenhouse gas, a reduction in oceanic carbon dioxide stores is an important consideration. UV-B-derived reductions in phytoplankton biomass are not presently incorporated into global climate change models.
Any sizable reduction in the population of nitrogen-fixing organisms may diminish the availability of nitrogen for higher plants (both naturally occurring species and crop plants). As a result, artificial nitrogen fertilization may be needed. However, such needs may stress the capabilities of developing countries.
A number of marine systems, particularly some fisheries (which are severely depleted due to over-harvesting and pollution), are presently stressed by anthropogenic factors. Coral reefs (important to both fisheries and tourism) are under stress and are experiencing declines from sedimentation, pollution, and perhaps temperature increases. Increased UV-B radiation may push some populations past their threshold by decreasing larval fish survival or increasing coral bleaching events.
Researchers continue to investigate UV-B penetration into the water column and through ice. Basic physical information could then be used in both laboratory and field research to determine their effects on phytoplankton and zooplankton. Adaptive strategies utilized by these communities, as well as the effects of interactive stresses (temperature, salinity, and cumulative doses received over the lifetime of the organisms, etc.) need to be studied over extended periods of time. As a result, plankton effects can be modeled into broader ecosystem dynamic patterns and can be fed into models of global carbon and nitrogen cycling.
Andreae, M.O., The ocean as a source of atmospheric sulphur compounds, pp. 331-362 in The Role of Air-Sea Exchange in Geochemical Cycling, P. Buat-Mernard (ed.), Deidel, Dordrecht, 1986.
Baker, K.S. and R.C. Smith, Spectral irradiance penetration in natural waters, pp. 223-246 in The Role of Solar Ultraviolet Radiation in Marine Ecosystems, J. Calkins, (ed.), Plenum, New York, 1982.
Bidigare, R.R., Potential effects of UV-B radiation on marine organisms of the Southern Ocean: distributions of phytoplankton and krill during Austral spring, Photochem. Photobiol., 50, 469 - 477, 1989.
Bittersmann, E., A.R. Holzwarth, G. Agel, and W. Nultsch, Picosecond time-resolved emission spectra of photoinhibited and photobleached Anabaena variabilis, Photochem Photobiol., 47, 101-105, 1988.
Carretto, J.J., M.O. Carignana, G. Daleo, and S.G. de Marco, Occurrence of mycosporine-like amino acids in the red tide dinoflagellate Alexandrium excavarum, UV-photoprotective compounds, J. Plankton Res., 12, 909-921, 1990.
Döhler, G., Effect of UV-B (290-320 nm) radiation on uptake of 15N-nitrate by marine diatoms, pp. 354-359 in Inorganic nitrogen in plants and microorganisms: Uptake and metabolism, W.R. Ullrich, C. Rigano, A. Fuggi, J.P. Aparicio, (eds.), Springer Verlag, Berlin, Heidelberg, New York, 1990.
Döhler, G. and M.R. Alt, Assimilation of 15N-ammonia during irradiance with ultraviolet-B and monochromatic light by Thalassiosira rotula, Compt. Rend., Acad. Sci. Paris, Serie D., 308, 513-518, 1989.
Eggersdorfer, B. and D.-P. Häder, Phototaxis, gravitaxis, and vertical migration in the marine dinoflaggelates, Peridinium faeroense and Amphidinium caterea, Acta Protozool., in press, 1991.
Garcia-Pichel, F. and R.W. Castenholz, Characterization and biological implications of scytonemin, a cyanobacterial sheath pigment, J. Phycol., 27, 395-409, 1991.
Garrison, D.L., C.W. Sullivan, and S.F. Ackley, Sea ice microbial communities in Antarctica, Bio. Science, 4, 243-250, 1986.
Gieskes, W.C. and G.W. Kraay, Transmission of ultraviolet light in the Weddell Sea: Report on the first measurements made in Antarctic, Biomass Newsletter, 12, 12-14, 1990.
Häder, D.-P. and S.M. Lui, Mobility and gravitactic orientation of the flagellate, Euglena gracilis, impaired by artificial and solar UV-B radiation, Curr. Microbiol., 21, 161-168, 1991.
Häder, D.-P. and R.C. Worrest, Effects of enhanced solar radiation on aquatic ecosystems, Photochem. Photobiol., 53, 717-725, 1991.
Holm-Hansen, O., UV radiation in Arctic waters: Effect on rates of primary production, Appendix G., pp. 1-17 in Proceedings of Workshop on "Response of Marine Phytoplankton on Natural Variation in UV-B Flux," Scripps Institution of Oceanography, May 1990.
Houghton, R.A. and G.M. Woodwell, Global climactic change, Sci. Amer., 260, 36-44, 1989.
Ignatides, L., Photosynthetic capacity of the surface microlayer during the mixing period, J. Planktonic Res., 12, 851-860, 1990.
IPCC Scientific Assessment, J.T. Houghton, G.J. Jenkins, and J.J. Ephraums (eds.), Cambridge University Press, Cambridge, 1991.
Jeffery, S.W. and G.H. Humprey, New spectrophotometric equations for determining chlorophylls a, b, c1, and c2 in higher plants, algae, and natural phytoplankton, Biochem. Physiol. Pflanzen, 167, 191-194, 1975.
Karentz, D., J.E. Cleaver, and D.L. Mitchell, DNA damage in the Antarctic, Nature, 350, 28, 1991a.
Karentz, D. and L.H. Lutze, Evaluation of biologically harmful ultraviolet radiation in Antarctica with a biological dosimeter designed for aquatic environments, Limnol. Oceanogr., 35, 549-561, 1990.
Karentz, D., F.S. Mc Euen, M.C. Land, and W.C. Dunlap, Survey of microsporine-like amino acid compounds in Antarctic marine organisms: potential protection from ultraviolet exposure, Marine Biology, 108, 157-166, 1991b.
Karsten, U., C. Wiencke, and G.O. Kirst, The effect of light intensity and daylength on the ß-dimethylsulphonio-propionate (DMSP) content of marine green macroalgae from Antarctica, Plant Cell Environm., 12, 989-993, 1990.
Kumar, A. and H.D. Kumar, Nitrogen fixation by blue-green algae, pp. 85-103 in Plant Physiology Research, S.P. Seu (ed.), Society for Plant Physiology and Biochemistry, 1st International Congress of Plant Physiology, New Dehli, India, Feb., 15-22, 1988.
Lubin, D., J.E. Fredrick, C.R. Booth, T. Lucas, and D. Neuschuler, Measurements of enhanced springtime ultraviolet radiation at Palmer Station, Antarctica, Geophys. Res. Lett., 16, 783-785, 1989.
Mitchell, B.G., Action spectra of ultraviolet photoinhibition of Antarctic phytoplankton and a model of spectral diffuse attenuation coefficients, Appendix G., pp. 1-15 in Proceedings of Workshop on "Responses of Marine Phytoplankton to Natural Variations in UV-B Flux," Scripps Institution of Oceanography, May 1990.
Nultsch, W., J. Pfan, and K. Huppertz, Photoinhibition of photosynthetic oxygen production and its recovery in the subtidal red data Polyneura hilliae, Bot. Acta, 103, 62-67, 1990.
Raven, J.A., Responses of aquatic photosynthetic organisms to increased solar UV-B, J. Photochem. Photobiol. B: Biol., 9, 239-244, 1991.
Smith, R.C., K.S. Baker, O. Holm-Hansen, and R. Olson, Photoinhibition of photosynthesis in natural waters, Photochem. Photobiol., 31, 585-592, 1980.
Vosjan, J.H., G. Döhler, and G. Nieuwland, Effect of UV-B irradiance on the ATP content of microorganisms of the Weddell Sea Antarctica, Neth. J. Sea Res., 25, 391-394, 1990.
Voytek, M.A., Addressing the biological effects of decreasing ozone in the Antarctic environment, Ambio, 19(2), 52-61, 1990.
Yoder, J.A. and S.S. Bishop, Effects of the mixing-induced irradiance fluctuations on photosynthesis of natural assemblages of coastal phytoplankton, Mar. Biol., 90, 87-93, 1990.
Zundorf, I. and D.-P. Häder, Biochemical and spectroscopic analysis of UV effects on the marine flagellate, Cryptomonas maculata, Arch. Microbiol., in press, 1991.
UNEP, Environmental Effects Panel Report, J.C. van der Leun and M. Tevini, (eds.), United Nations Environment Programme, Nairobi, Kenya, 1989.
{kind=link}
{kind=link}
{kind=link}